ќдн≥Їю з найпоширен≥ших груп безхребетних, ¤к≥ використовують дл¤ палеогеограф≥чних реконструкц≥й, Ї молюски; це справедливо й стосовно палеоб≥огеох≥м≥чних досл≥джень. «а скромними п≥драхунками понад 2/3 палеоб≥огеох≥м≥чних праць ірунтуютьс¤ на вивченн≥ черепашок б≥вальв≥й ≥ гастропод. ÷е зумовлено низкою причин, важливими з ¤ких Ї: значне поширенн¤ представник≥в цих клас≥в у —в≥товому океан≥ та, особливо, в еп≥континентальних мор¤х; а також здатн≥сть ≥снувати в водах з р≥зною солон≥стю практично у вс≥х кл≥матичних зонах. ћолюски досить поширен≥ в узбережних м≥лководних морських обстановках на глибинах, ¤к≥ не перевищують 150-200 м, де часто дом≥нують в донних б≥оценозах.
ћолюски, ¤к ≥ ≥нш≥ високоорган≥зован≥ групи безхребетних, утворюють своњ скелети переважно секрец≥йним способом, мають м≥нерально-орган≥чну черепашку, досить ст≥йку у п≥сл¤седиментац≥йних умовах, що забезпечуЇ збереженн¤ первинного прижиттЇвого складу скелетноњ речовини. ‘ормуЇтьс¤ черепашка молюск≥в з екстрапал≥альноњ р≥дини, головн≥ компоненти ¤коњ зумовлен≥ складом гемол≥мфи та через нењ ф≥зико-х≥м≥чними особливост¤ми вод проживанн¤. «авд¤ки досить ст≥йк≥й залежност≥ черепашка-екстрапал≥альна р≥дина-гемол≥мфа-води проживанн¤ б≥огеох≥м≥чн≥ показники можна використати дл¤ реконструкц≥њ умов середовища. “ривалий час припускали, що склад екстрапал≥альноњ р≥дини та вод проживанн¤ абсолютно ≥дентичн≥ [9], сьогодн≥ очевидно, що це не зовс≥м так.
≈кспериментальн≥ дан≥ засв≥дчили, що черепашка молюска формуЇтьс¤ в х≥м≥чн≥й р≥вноваз≥ з водами проживанн¤ далеко не завжди [8]. ™ ф≥з≥олог≥чн≥ механ≥зми, ¤к≥ регулюють надходженн¤ х≥м≥чних елемент≥в з зовн≥шнього середовища до внутр≥шн≥х та естрапал≥альних флюњд≥в. ÷е, з одного боку, значно ускладнюЇ процедуру в≥дновленн¤ умов середовища проживанн¤ палеоб≥огеох≥м≥чними методами, та, з ≥ншого - даЇ змогу одержати повн≥шу незалежну ≥нформац≥ю, залучаючи до об'Їкт≥в вивченн¤ види, ¤к≥ мають р≥зн≥ апарати ф≥з≥олог≥чних адаптац≥й.
Ќайв≥дом≥ш≥ реконструкц≥њ за палеоб≥огеох≥м≥чними даними - це оц≥нки солоност≥ та температури вод проживанн¤. Ќа п≥дстав≥ таких досл≥джень робили спроби в≥дновити ф≥зико-х≥м≥чн≥ умови морських басейн≥в крейдового пер≥оду ≥ кайнозою на значних за площею територ≥¤х [7]. ¬одночас далеко не завжди палеоб≥огеох≥м≥чн≥ ≥нформац≥¤ про температуру та солон≥сть палеобасейн≥в п≥дтверджувана ≥ншими незалежними методами. «вичайно так≥ в≥дхиленн¤ по¤снюють впливом п≥сл¤седиментац≥йних зм≥н черепашковоњ речовини. Ќе в≥дкидаючи цього, зазначимо, що в багатьох випадках причиною недостатньоњ ефективност≥ палеоб≥огеох≥м≥чних методик Ї не п≥сл¤седиментац≥йн≥ перетворенн¤, а неврахуванн¤ п≥д час ≥нтерпретац≥њ б≥огеох≥м≥чних даних ф≥з≥олог≥чних особливостей молюск≥в. Ќаш≥ досл≥дженн¤ довели, що одним з найд≥Їв≥ших механ≥зм≥в, ¤кий визначаЇ режим формуванн¤ черепашки молюск≥в, Ї осморегулюванн¤.
ћолюски, ¤к ≥ б≥льш≥сть ≥нших тварин, ¤к≥ проживають у водному середовищ≥, повинн≥ захищати р≥динн≥ середовища свого орган≥зму в≥д впливу екстремальних осмотичних умов довк≥лл¤ [6]. –≥зн≥ групи молюск≥в мають далеко не однаков≥ механ≥зми осморегулюванн¤, що в≥дображено й на склад≥ скелетноњ речовини, ¤ка утворюютьс¤ з екстрапал≥альноњ р≥дини. «а нашими даними, що одержан≥ п≥д час вивченн¤ ¤к викопних, так ≥ сучасних водних молюск≥в, спос≥б осморегулюванн¤ Ї одним з головних чинник≥в, ¤кий зумовлюЇ ефективн≥сть палеоб≥огеох≥м≥чного анал≥зу.
Ѕ≥льш≥сть жител≥в морських вод Ї осмоконформаторами, тобто тон≥чн≥сть њхнього внутр≥шнього середовища (гемол≥мфи та екстрапал≥альноњ р≥дини) приблизно в≥дпов≥даЇ тон≥чност≥ навколишн≥х вод. ѕовн≥стю це справджуЇтьс¤ стосовно стеногал≥нних форм, ¤к≥ здатн≥ нормально розвиватис¤ у водах пор≥вн¤но вузького д≥апазону солоност≥. «а умов п≥двищенн¤ або зниженн¤ солоност≥ вод стеногал≥нн≥ молюски ризикують загинути в≥д осмотичного збезводненн¤ або обводненн¤. ” вузькому д≥апазон≥ зм≥н солоност≥ захист в≥д небажаних ¤вищ забезпечують засоби внутр≥шньокл≥тинноњ ≥зоосмот≥њ [9], ¤ка регулюЇ тон≥чн≥сть цитоплазми. ¬нутр≥шньокл≥тинна ≥зоосмот≥¤ реал≥зована за допомогою двох механ≥зм≥в, робота ¤ких ч≥тко координована на тл≥ зовн≥шн≥х умов, ¤к≥ зм≥нюютьс¤. ќдин з цих механ≥зм≥в пов'¤заний з≥ зм≥нами концентрац≥њ у цитоплазм≥ орган≥чних осмол≥т≥в, ≥нший ірунтуЇтьс¤ на транспортуванн≥ через кл≥тинн≥ мембрани неорган≥чних ≥он≥в. јктивне транспортуванн¤ ≥он≥в регульоване спец≥альними ферментами та в≥дбуваЇтьс¤ з витратами енерг≥њ. «м≥на концентрац≥њ орган≥чних з'Їднань економн≥ша тому, що орган≥чн≥ молекули не проход¤ть через кл≥тинн≥ мембрани ≥ не витрачаютьс¤ в процес≥ регулюванн¤ тон≥чност≥.
≈вригал≥нн≥ морськ≥ молюски також мають здатн≥сть до ≥зоосмотичного регулюванн¤ внутр≥шньокл≥тинних флюњд≥в, проте вони можуть у пор≥вн¤но вузькому д≥апазон≥ солоност≥ вод регулювати тон≥чн≥сть позакл≥тинних р≥дин. “ому в ¤комусь д≥апазон≥ солоност≥ позакл≥тинн≥ р≥дини евригал≥нних молюск≥в збер≥гають в≥дносну незалежн≥сть своЇњ тон≥чност≥, завд¤ки чому ≥зоосмот≥¤ тканин пов'¤зана з подоланн¤м дещо меншого град≥Їнта тиску [6].
«≥ сказаного випливаЇ, що молюски, ¤к≥ належать до стеногал≥нноњ та евригал≥нноњ груп, мають р≥зн≥ механ≥зми в≥дображенн¤ умов середовища у своњх позакл≥тинних р≥динах та, ¤к насл≥док, у скелет≥, ¤кий утворюЇтьс¤ з одн≥Їњ з цих р≥дин. ” стеногал≥нних молюск≥в склад екстрапал≥альноњ р≥дини зм≥нюЇтьс¤ грубо синхронно з коливанн¤м солоност≥ вод проживанн¤. ќчевидно, тому вм≥ст натр≥ю ≥ стронц≥ю в черепашках саме цих молюск≥в ≥нформуЇ про солон≥сть водного середовища найефективн≥ше [1].
ƒоведемо останнЇ припущенн¤ на приклад≥ багатих п≥зньобаденських малакоценоз≥в ¬олино-ѕод≥лл¤ [1]. як св≥дчить вм≥ст натр≥ю та стронц≥ю [5] - найефективн≥ших показник≥в палеосолоност≥, у черепашках б≥вальв≥й ≥ гастропод з нормально солоних та дещо опр≥снених зон (р≥зниц¤ оц≥нена у 3-5Й), далеко не вс≥ молюски однаково реагують на зм≥ни солоност≥ вод проживанн¤. « досить велико≥ к≥лькост≥ вивчених б≥вальв≥й ≥ гастропод лише Glycymeridae, Carditidae та де¤к≥ Trochidae ви¤вили себе ¤к ≥ндикатори солоност≥ вод. —учасн≥ представники роду Glycymeris живуть у водах з нормальною солон≥стю, кардитид теж уважають стеногал≥нною групою, ¤ка в≥ддаЇ перевагу нормальн≥й солоност≥ [4]. ≤ншою рисою, ¤ка об'ЇднуЇ згадан≥ родини, Ї те, що вони проживають на поверхн≥ ірунту або незначно заглиблюютьс¤ в нього. « рисунку 1, ¤кий ≥люструЇ розпод≥л натр≥ю та стронц≥ю у черепашках молюск≥в з п≥дг≥рських та терноп≥льських верств верхнього баден≥ю, видно, що з семи протестованих вид≥в т≥льки три накопичували Na ≥ Sr у р≥вноваз≥ з солон≥стю вод проживанн¤; це б≥вальв≥њ Glycymeris pilosa deshayesi (May) ≥ Cardites rudista (Desh.) - представники згаданих вище гл≥цимер≥д и кардитид, та гастроподи Gibbula buchi (Dub.) [2]. —аме ц≥ три види Ї типово стеногал≥нними, тод≥ ¤к представники ≥нших чотирьох вид≥в менше чутлив≥ до зм≥н солоност≥ вод проживанн¤. Ћ≥воруч на рис. 1 показана концентрац≥¤ водорозчинного натр≥ю у черепашках устриць, ¤ка досить над≥йно в≥дображаЇ коливанн¤ солоност≥ вод проживанн¤ [1] й використана нами ¤к незалежний показник.
¬нутр≥шн≥ флюњди орган≥зму пр≥сноводних тварин, в т≥м числ≥ й молюск≥в, звичайно г≥перосмотичн≥ пор≥вн¤но з њхн≥м водним оточенн¤м. «деб≥льшого пр≥сноводн≥ молюски Ї осморегул¤торами. ќсморегулюванн¤ у них в≥дбуваЇтьс¤ завд¤ки п≥двищенню тон≥чност≥ внутр≥шнього середовища та вид≥ленн¤ б≥льшоњ к≥лькост≥ г≥потон≥чноњ сеч≥. ƒеф≥цит сольового балансу, ¤кий виникаЇ п≥д час п≥дтриманн¤ такого режиму, покриваЇ активна сорбц≥¤ ≥он≥в ззовн≥. ÷е в≥дбуваЇтьс¤ проти осмотичного град≥Їнта й тому супроводжуЇтьс¤ затрачанн¤м енерг≥њ. ќтже, екстрапал≥альна р≥дина, ¤к ≥ ≥нш≥ позакл≥тинн≥ флюњди пр≥сноводних молюск≥в, значно б≥льше збагачена сол¤ми, н≥ж навколишн¤ вода. “ому й вм≥ст у черепашках таких визнаних ≥ндикатор≥в солоност≥, ¤к натр≥й та стронц≥й, навр¤д чи успадковуЇ м≥нерал≥зац≥ю вод.
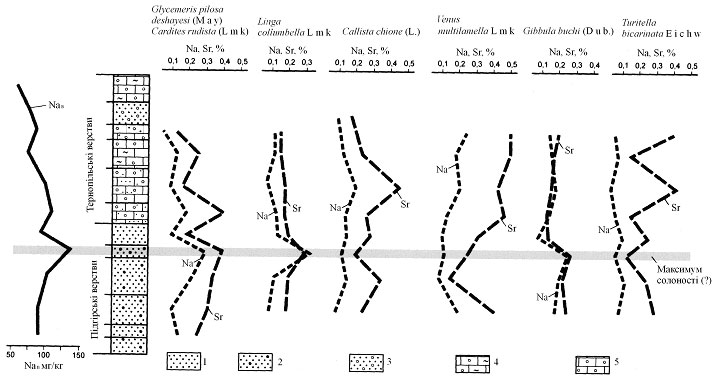
–ис.1. ¬м≥ст натр≥ю та стронц≥ю в черепашках стеногал≥нних та евригал≥нних молюск≥в з середньо м≥оценових в≥дклад≥в ¬олино- ѕод≥лл¤. Ћ≥воруч показан≥ вар≥ац≥њ вм≥сту водорозчинного натр≥ю у черепашках Ostrea edulis digitalina Dub. з тих же в≥дклад≥в:
1 - п≥сок; 2 - п≥сок з дом≥шкою грав≥ю; 3 - п≥сок з жовнами л≥тотам н≥й; 4 - глинистий вапн¤к з л≥тотамн≥¤ми; 5 - л≥тотамн≥Ївий вапн¤к
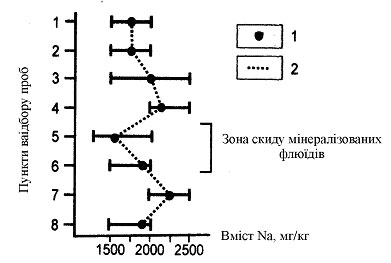
” черепашках Unio tumidus (Phil.) з р. ¬орскли в зон≥ надходженн¤ у р≥чков≥ води хлоридно-натр≥Ївих м≥нерал≥зованих пластових флюњд≥в [3], що виникають унасл≥док розробки родовищ вуглеводн≥в, п≥двищенн¤ вм≥сту натр≥ю не заф≥ксовано, кр≥м того, дл¤ цього х≥м≥чного елемента в зонах техногенного забрудненн¤ ви¤влене зменшенн¤ концентрац≥њ (рис. 2). ÷ей ефект, в≥рог≥дно, пов'¤заний з виникненн¤м у б≥отоп≥ стресовоњ ситуац≥њ, а визначений цей зв'¤зок також процесами осморегулюванн¤.
ѕр≥сноводн≥ молюски мають ф≥льтрац≥йно-реабсорбц≥йну осморегул¤ц≥йну систему, завд¤ки ¤к≥й поверненн¤ в орган≥зм з ф≥льтрату ф≥з≥олог≥чно ц≥нних речовин, у т≥м числ≥ ≥он≥в Na+, K+, Cl-, Ca2+, в≥дбуваЇтьс¤ через реабсорбц≥ю. “оксичн≥ або непотр≥бн≥ дл¤ орган≥зму речовини виход¤ть з сечею [6]. «а на¤вност≥ такого механ≥зму нема потреби активно переносити до сеч≥ токсичн≥ компоненти. ” випадку високого вм≥сту токсичних речовин у ф≥льтрат≥ осморегул¤ц≥йна система втрачаЇ ефективн≥сть, унасл≥док чого у позакл≥тинн≥ р≥дини все ж потрапл¤Ї де¤ка частина шк≥дливих компонент≥в та зменшуЇтьс¤ поглиненн¤ необх≥дних солей. «рештою, найб≥льше ц≥ в≥дхиленн¤ в≥д норми трапл¤ютьс¤ у склад≥ екстрапал≥альноњ р≥дини, з ¤коњ й формуЇтьс¤ черепашка з п≥двищеним вм≥стом токсичних елемент≥в та деф≥цитом натр≥ю. ” кристал≥чн≥й гратц≥ арагон≥ту м≥сце натр≥ю на р≥зних позиц≥¤х займаЇ стронц≥й (див. рис. 2), залученн¤ ¤кого до нењ за ≥нших однакових зовн≥шн≥х умов Ї виг≥дн≥шим енергетично.
ќтже, у раз≥ використанн¤ б≥огеох≥м≥чних показник≥в з метою палеогеограф≥чних реконструкц≥й умов минулого потр≥бно враховувати ф≥з≥олог≥чн≥ особливост≥ орган≥зм≥в-≥ндикатор≥в, ¤к≥ застосовують.
ƒл¤ оц≥нок солоност≥ вод проживанн¤ л≥пше застосовувати стеногал≥нн≥ форми молюск≥в, ¤к≥ практично не здатн≥ регулювати склад своњх внутр≥шньомант≥йних р≥дин, завд¤ки чому њхн≥й екстрапал≥альний флюњд майже не в≥др≥зн¤Їтьс¤ за х≥м≥чним складом в≥д вод проживанн¤. ћожна передбачити, що в цьому випадку вар≥ац≥њ вм≥сту натр≥ю ≥ стронц≥ю у черепашках будуть найточн≥ше в≥дображати зм≥ни солоност≥ вод. ѕевна р≥ч,
оч≥кувати повноњ ≥дентичност≥ цих показник≥в не доводитьс¤, бо формуванн¤ компонент≥в черепашки в≥дбуваЇтьс¤ в м≥кроумовах.
–ис. 2. ¬м≥ст натр≥ю в черепашках Unio tumidus з р. ¬орскли:
1 - середнЇ значенн¤ вм≥сту натр≥ю; 2 - розмах значень. Ќомери пункт≥в в≥дбору проб зростають за теч≥Їю р≥чки.
≈вригал≥нн≥ форми здатн≥ регулювати тон≥чн≥сть своњх мант≥йних р≥дин у певному д≥апазон≥ солоност≥, тому њхн¤ гемол≥мфа та екстрапал≥альна р≥дина, ¤ка утворюютьс¤ з гемол≥мфи, можуть збер≥гати в цьому д≥апазон≥ св≥й склад незалежно в≥д зовн≥шнього середовища. ≈вригал≥нн≥ молюски також зм≥нюють склад своњх черепашок залежно в≥д солоност≥ вод проживанн¤, однак, вони Ї менш точним ≥нструментом пор≥вн¤но з≥ стеногал≥нними формами.
ѕр≥сноводн≥ молюски належать до типових осморегул¤тор≥в, тому використанн¤ традиц≥йних б≥огеох≥м≥чних показник≥в дл¤ оц≥нок солоност≥ вод проживанн¤ в цьому випадку неефективне.
Ћ≥тература
Ѕорисенко ё.ј., ¬асильев ј.Ќ. Ѕиогеохими¤ раковин ископаемых двустворок как индикатор палеосолености // ѕалеонтологические и биостратиграфические исследовани¤ при геологической съемке на ”краине. - .: Ќаук. думка, 1990. - —. 120-126.
¬асильев ј.Ќ. —оленость вод позднебаденского бассейна ¬олыно-ѕодолии по палеобиогеохимическим данным // ≈косистеми геолог≥чного минулого ”крањни. - .: ≤√Ќ ЌјЌ ”крањни, 1995. - —. 97-98.
¬асильев ј.Ќ., ∆уравель Ќ.≈., лочко ѕ.¬. ќрганизаци¤ гидрохимического мониторинга в услови¤х нефтегазоносного северо-востока ”краины. - ’.: Ёкограф, 2001. - 112 с.
удрин Ћ.Ќ. —тратиграфи¤, фации и экологический анализ фауны палеогеновых и неогеновых отложений ѕредкарпать¤. - Ћ.: »зд-во Ћьвов. ун-та, 1966. - 174 с.
Ќевесска¤ Ћ.ј., √ончарова ».ј., »льина Ћ.Ѕ. и др. »стори¤ неогеновых моллюсков ѕаратетиса - ћ.: Ќаука, 1986. - 208 с.
Ёккерт –., –энделл ƒ., ќгастин ƒж. ‘изиологи¤ животных: ћеханизмы и адаптаци¤. - ћ.: ћир, 1992. - 344 с.
ясаманов Ќ.ј. ƒревние климаты «емли. - Ћ.: √идрометеоиздат, 1985. - 295 с.
Buchardt B., Fritz P. Strontium uptake in shall aragonite from the freshwater gastropod Limnaea stagnalis // Science.- 1978. - Vol. 199. - –. 291-292.
Dodd J.R. Magnesium and strontium in calcareous skeletons: а review // J. Paleontol.- 1967. - Vol. 41. - P. 1313-1329.
Schmidt-Nielson K. Countercurrent system in animals // Scientific American. - 1984. - Vol. 224.- –. 118-128.