Територія Подільського Придністер'я багата на палеолітичні пам'ятки середнього і пізнього палеоліту. Зокрема, тут розташовані європейського значення стоянки Молодове І і V, Кормань ІV, Кертоси, Стінка та ін. Окремі з них відомі ще з першої третини (Молодове І), інші з середини ХХ століття. На цих стоянках працювали такі відомі археологи, як О. Черниш і М. Анісюткін, геологи і палеогеографи І. Іванова, В. Мотуз, С. Губін, Н. Ренгартен, Н. Боліховська, Г. Пашкевич та ін. [3, 4, 6, 8-10]. У 70-80-х роках ХХ ст. вийшла серія монографій, присвячених окремим стоянкам, палеогеографічним умовам проживання на них давньої людини. Питанню екології палеолітичної людини присвячені також праці І. Іванової [5].
Артефакти на згаданих палеолітичних стоянках залягають у відкладах лесово-ґрунтової серії, яка часто має сліди делювіального перевідкладення. Потужність їх, залежно від розташування стоянки, коливається у межах 3-10 м, а подекуди й більше. Оскільки ці відклади є практично повсюдно карбонатними, то вони містять значну кількість фауни молюсків. Аналіз цієї фауни дав змогу І. Івановій та В. Мотузу відтворити окремі риси палеогеографічної обстановки часу формування лесово-ґрунтової серії і, відповідно, палеолітичної людини.
Упродовж 1998-1999 рр. міжнародна археологічно-геологічна експедиція провадила розкопки на палеолітичній стоянці Молодове V, розташованій на правому березі Дністра поблизу с. Братанівка Сокирянського р--у Чернівецької обл. (рис. 1). До комплексу виконуваних робіт належали і малакологічні. Дослідники мали на меті порівняти отриману фауну з результатами попередніх досліджень, а також відтворити природні умови формування досліджуваної пачки відкладів. Ми розглянемо фауну молюсків, виявлену у відкладах з мустьєрськими палеолітичними горизонтами, які відповідають у часовому зрізі першій половині в'юрму.
Викопну фауну молюсків району палеолітичної стоянки Молодове V висвітлено у працях І. Іванової і В. Мотуза [9]. В. Мотуз на підставі проб, відібраних у ході археологічних робіт експедицією під керівництвом О. Черниша, детально проаналізовано викопну малакофауну окремих стратиграфічних горизонтів. Дослідник опробував усю товщу відкладів, які вкривають другу надзаплавну терасу Дністра - від еему до голоцену, визначив близько 50 видів та підвидів викопних гастропод. Він зазначив, що у період формування мустьєрських культурних шарів (12-10а за схемою О. Черниша [9] ) у фауні домінували убіквісти - види, які проживають у широких за діапазоном температур та вологості умовах. Це, зокрема, такі види, як Succinea oblonga elongata Sandb. i Pupilla muscorum L. Також він виявив, що керівними для цих горизонтів були холодолюбні (кріофільні) види, такі як Columella columella Mart. і Vallonia tenuilabris Al.Br., що свідчить про холодні умови формування лесово-ґрунтової товщі. Щодо ценозів, які сформувалися в час нагромадження товщі, то визна-чальними для них можна вважати такі види: Pupilla muscorum L., Pupilla sterri Voith., Vallonia pulchella Mull., Vallonia costata Mull., Vallonia tenuilabris Al.Br. - індикатори відкритих просторів, Columella edentula Drap., Clausilia dubia Drap., Eucunulus fulvus Mull. та інші, які проживали в затінених місцях. Кліматичні умови, у яких відкладалася лесова товща, за даними В. Мотуза, часто і різко змінювалися під впливом неодноразових похолодань і потеплінь упродовж останнього зледеніння.
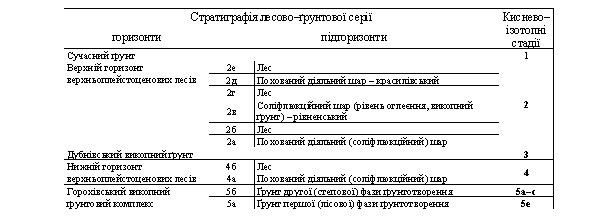
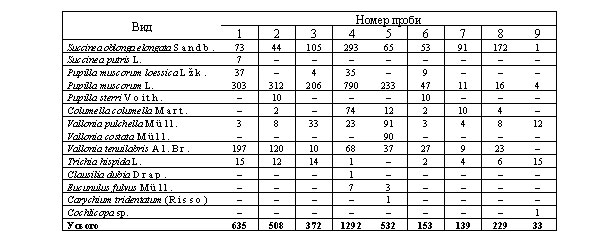
Під час малакофауністичних досліджень ми відібрали 18 проб з усіх горизонтів, виділених геологом А. Богуцьким. Для опису розрізу (рис. 2) використали стратиграфічну схему, приведену у табл. 1 [1, 12]. До мустьєрських шарів належать зразки з першого по дев'ятий, склад яких наведено у табл. 2. Інтерпретацію малакофауни виконано за допомогою праць [2, 7, 11, 13].
Рельєф стоянки (розташування на схилі другої тераси Дністра [9]) не виключає переміщення матеріалу вниз по схилу і, відповідно, його змішування. Тому треба враховувати імовірність цих процесів протягом усього часу формування лесово-ґрунтової пачки - можливе домішування видів, які були знесені з вищих ділянок долини.
Таблиця 1
Горизонти і підгоризонти верхнього плейстоцену Волино-Поділля [12]
Таблиця 2
Фауна молюсків мустьєрських культурних шарів палеолітичної стоянки Молодове V
Отримана фауна молюсків дає змогу зробити такі палеокліматичні інтерпретації щодо часу формування досліджуваної товщі.
Головними видами, які утворюють малакологічний комплекс проби 1, є Pupilla muscorum L. (47,7%) і Vallonia tenuilabris Al.Br. (31,0%). Значну частину фауни становлять холодолюбні види. До них належить Vallonia tenuilabris Al.Br., Pupilla loessica Lzk., їхній сумарний відсоток сягає 36,8. Вид Pupilla loessica Lzk. свідчить про процес еолової акумуляції лесового матеріалу. Наявність видів Succinea oblonga elongata Sandb. і, особливо, Succinea putris L. (сумарна частка їх становить 12,6) є ознакою досить значної зволоженості території. Вся фауна засвідчує, що на той час характерними для досліджуваної території були відкриті холодні біоценози, можливо, так звані тундролуки і тундростепи.
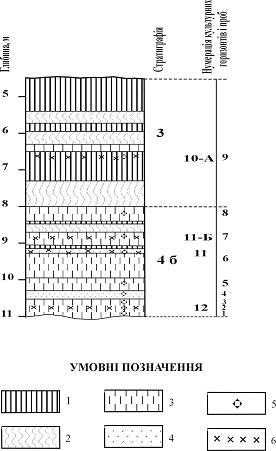
Рис. 2. Розріз палеолітичної стоянки Молодове V:
1 - горизонт А1 викопних ґрунтів; 2 - горизонт В викопних ґрунтів; 3 - супіски; 4 - глейові горизонти; 5 - проби, у яких виявлено фауну молюсків; 6 - культурні горизонти.
У пробі 2, як і в попередній, комплекс ви-копних молюсків формують два види - Pupilla muscoru L. (63,4%) і Vallonia tenuilabri Al.Br. (23,6%). У цьому зразку зникає вид Pupilla loessica Lzk., але з'явля-ється, хоча у дуже малій кількості, Columella colu-mella Mart. Загальне змен-шення частки холодо-любних форм свідчить, імовірно, про незначне, порівняно з попереднім зразком потепління. У цій пробі цікавий також ще один новий вид - Pupilla sterri Voith. Його поява, а також зменшення кількості Succinea oblonga elonga-ta Sandb. і зникнення Succinea putris L. є ознакою арідизації клімату. Склад усього малакологічного комплексу засвідчує, що на досліджуваній території, як і в попередньому випадку, переважала рослинність від-критих незаліснених біо-топів. Цій пробі відповідає 12-й культурний шар. Проба 3 відібрана з товщі 0,15-0,20 м над 12-м культурним шаром.
Головними видами, які формують комплекс є Pupilla muscorum L. (55,4 %) і Succinea oblonga elongata Sandb. (28,2 %). Характер фауни - зменшення (лише 3,8 %) кількості холодолюбних видів - відображає незначне потепління клімату. Поява виду Pupilla loessica Lzk. свідчить про відновлення процесу лесової акумуляції. На підставі збільшення частки виду Succinea oblonga elongata Sandb. можна зробити висновок про зростання зволоженості на досліджуваній території в той час.
Як і в попередньому випадку, у пробі 4 панівними видами залишилися Pupilla muscorum L. і Succinea oblonga elongata Sandb. - відповідно, 61,1 і 22,7%. Цей етап характерний тим, що значно збільшилася частка в якісному та кількісному аспектах холодолюбної аркто-бореальної фауни: до згадуваних у попередній пробі двох видів додався ще Columella columella Mart. - їхня загальна кількість становила 13,6%.
Зростання кількості особин виду Pupilla loessica Lzk. свідчить про посилення процесу накопичення лесового пилу. Для цієї проби характерне зменшення, порівняно з іншими пробами, кількості особин виду Trichia hispida L., що також є ознакою похолодання та ймовірної аридизації. Доказом цього також, можливо, є зменшення, хоча і незначне, кількості особин Succinea oblonga elongata Sandb. Треба зазначити про появу форм, які раніше не траплялися - Clausilia dubia Drap. (0,1%) і Eucunulus fulvus Mull. (0,5%). Вони є типовими тіньолюбними видами. Тобто на цій території в той час переважала трав'яниста рослинність, хоча, ймовірно, у невеликій кількості траплялися дерево-чагарникові рослинні угруповання.
Фауна зразка 5 змінилася порівняно з попередньою пробою. Найбільшу частку проби займає, як і раніше, Pupilla muscorum L. На другій позиції опинилися зразу три види, відсоток яких у пробі коливається в межах 12-17 - це Succinea oblonga elongata Sandb., Vallonia pulchella Mull., а також новий таксон Vallonia costata Mull. Зменшення кількості екземплярів Succinea oblonga elongata Sandb., зростання Vallonia pulchella Mull. та поява oblonga elongata Sandb., Vallonia pulchella Mull., а також новий таксон Vallonia costata Mull., який тяжіє до ксерофілів, свідчить про загальну аридизацію клімату. Порівняно з попереднім етапом зменшилася кількість кріофільної фауни: взагалі зник Pupilla loessica Lzk., зменшився відсоток Columella columella Mart., хоча і збільшилася кількість особин Vallonia tenuilabris Al.Br. Для описуваного горизонту характерні також тіньолюбні види - Eucunulus fulvus Mull. (0,6%) і Carychium tridentatum (Risso) (0,2%). Отже, в той час відбулося незначне потепління, яке супроводжувалося загальним осушенням клімату, панівними біоценозами залишилися тундролукостепові. Характерною рисою цього горизонту є знаходження залишків 11-го культурного шару .
На підставі аналізу решток викопного малакоценозу проби 6, можна сказати таке: відбулося чергове похолодання. Про це свідчить збільшення холодолюбної фауни - знову з'явився Pupilla loessica Lzk. (5,9%), суттєво збільшилася кількість Vallonia tenuilabris Al.Br. (17,6%), однак дещо зменшилася кількість Columella columella Mart. (1,3%). Наявність Pupilla loessica Lzk. свідчить про відновлення процесу лесової акумуляції. Збільшення кількості особин виду Succinea oblonga elongata Sandb. є ознакою зростання зволоженості. Водночас знову з'явився Pupilla sterri Voith. (6 %) - типовий ксерофільний вид. Характерним є зникнення будь-яких тіньолюбних таксонів, що засвідчує можливу заміну рослинних угруповань тундростеповими.
Проба 7 відібрана з типових лесів, які залягають на 0,3 м вище від горілого горизонту, який вміщує культурний шар 11-Б. З цієї проби починається суттєве зменшення кількості особин виду Pupilla muscorum L. Домінує тут Succinea oblonga elongata Sandb., кількість якого становить 69,5%. Значне збільшення кількості особин цього виду, а також збільшення відсотка вологолюбного кріофіла Columella columella Mart. (7,6%) свідчать про зростання зволоженості території. Індикатором цього є також зникнення будь-яких ксерофільних видів. Тобто ценози, які панували в той час на описуваній території, були відкриті, холодні, досить добре зволожені (ймовірно, вологий тундролукостеп).
Як і в попередньому випадку, панівним видом у пробі 8 є Succinea oblonga elongata Sandb., кількість якого тут є максимальна для всього лесово-ґрунтового профілю Молодове V і становить 75,1%. Дещо зменшується загальний відсоток кріофілів зі зменшенням Columella columella Mart. (1,75%) та зростанням Vallonia tenuilabris Al.Br. (10,1%). Аналіз комплексу малакофауни свідчить про формування подібних до описуваних у попередній пробі біоценозів, хоча, швидше за все, вони були дещо теплішими.
Малакофауна зразка 9 суттєво відрізняється від усіх описаних вище. Зазначимо, що вона відібрана з викопного ґрунту, вік якого за І. Івановою [10] датовано як хенґело-подґрадем. У цьому викопному ґрунті знайдено останній (наймолодший) з мустьєрських культурних шарів - 10-А. Характерна риса проби - повна відсутність холодолюбних видів, що свідчить про теплі умови. Головними складовими цього малакологічного комплексу є Trichia hispida L. (46,9%) і Vallonia pulchella Mull. (37,5%), які й визначили умови формування цієї товщі. Малакофауністичні комплекси, у складі яких домінує Trichia hispida L., є ознакою порівняно теплих та вологих кліматичних умов. На підставі аналізу цієї проби можна сказати, що її фауна відповідає найтеплішому з усіх описаних вище етапів. У цей час на території в рослинному покриві домінували відкриті ценози, однак значно тепліші від попередніх, - імовірно, прирічкові луки.
Отже, природна обстановка протягом формування лесово-ґрунтової серії змінювалася: відбувалось послідовне чергування потеплінь та похолодань, з якими, відповідно, були пов'язані етапи зростання зволоженості або аридизації (осушення). Найтеплішим з описаних вище був період, під час якого сформувався дубнівський викопний ґрунт (у І. Іванової - хенґело-подґрадем); упродовж нього виник малакокомплекс, провідним видом у якому був Trichia hispida L.
Література
Богуцкий А.Б. Антропогеновые покровные отложения Волыно-Подолии // Антропогеновые отложения Украины. - Киев: Наук. думка, 1986. - С. 121-132.
Жадин В.И. Моллюски пресных солоноватых вод СССР. - М; Л.: Изд-во АН СССР, 1952. - 346 с.
Иванова И.К. Геологическое строение палеолитической стоянки Атаки 1 в Среднем Приднестровье // Бюлл. Комиссии по изуч. четвертич. периода АН СССР.- 1968. - №35. - С.113 - 119.
Иванова И.К. Геологическое строение долины р. Днестр в районе мустьерского местонахождения Стинка // Бюлл. Комиссии по изуч. четвертич. периода АН СССР. - 1968. - №36. - С. 129-136.
Иванова И.К. Природные условия обитания людей каменного века в басейне р. Днестр // Палеогеография древнего человека. - М.: Наука, 1977. - С. 7-18.
Кетросы. Мустьерская стоянка на Среднем Днестре. - М.: Наука, 1981. - С. 93-103.
Лихарев И.М., Раммельмейер Е.С. Наземные моллюски фауны СССР. - М.: Изд-во АН СССР, 1952. - 488 с.
Многослойная палеолитическая стоянка Кормань IV на Среднем Днестре.К X Конгрессу ИНКВА. - М.: Наука, 1977. - С.119 - 125.
Многослойная палеолитическая стоянка Молодова V. Люди каменного века и окружающая среда. К VII Конгрессу ИНКВА. - М.: Наука, 1982. - 239с.
Молодова І. Уникальное мустьерское поселение на Среднем Днестре. - М.: Наука, 1982. - С. 120-145.
Alexndrowicz S.W. Analiza malakologiczna w badaniach osdow czwartorzedowych // Geologia. - 1987. - T.13. - Z. 1-2. - S. 3-240.
Boguckyj A., Lanczont M. Syratygrafia lessow Naddniestra halickego // Studia Geologica Polonica. Lessy i paleolit Naddniestrza Halickego (Ukraina). - 2002. - Vol. 119. - S. 315-327.
Lozek V. Quartarmollusken der Tschechoslovakei. - Praha, 1964. - 374 s.